В дальнейшем селекционно-генетическая работа с гибридной пшеницей была направлена нами на создание и изучение самофертильной аллоцитоплазматической пшеницы (яровой и озимой), у которой геном T.aestivum L. нормально без нарушений функционирует в чужеродной цитоплазме, что подтверждается отсутствием у них явления ЦМС.
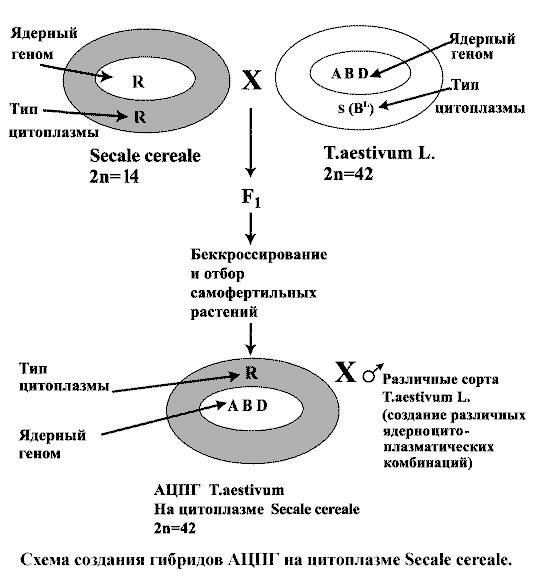
Исходные формы (яровые и озимые) аллоцитоплазматической пшеницы T. aestivum L. были созданы путем последовательного замещения ядра донора цитоплазмы (Aegilops ovata, T. timopheevi, Secale cereale L., и др.) на ядро пшеницы мягкой (T. aestivum L.) методом беккроссирования (не менее В6).
При этом как в первоначальных, так и в последующих скрещиваниях доноры цитоплазмы были использованы в качестве материнских форм (Медведева, Семенов, 1969, 1972). В процессе насыщающих обратных скрещиваний осуществляли постоянно отбор растений на самофертильность путем индивидуальной изоляции, а также по морфологическим признакам, характерным для сортов T. aestivum L. - доноров ядра. Эти гибридные формы были использованы в качестве доноров цитоплазмы при скрещивании их с коллекционными сортами.
Аллоцитоплазматические гибриды (АЦПГ) пшеницы мягкой представляют собой новый синтетический тип злаковых растений, у которых ядро T. aestivum L. нормально (без ЦМС) функционирует в чужеродной цитоплазме. Плазмо-ядерное взаимодействие у созданных форм индуцирует комплекс ценных признаков, сохраняющихся в поколениях. Это важно для селекционной работы, поскольку способствует обогащению генофонда этой культуры (Семенов, 1978).
Линии аллоцитоплазматической пшеницы являются удобным модельным объектом для изучения характера функциональных взаимодействий плазмагенов с хромосомной генной системой и для регулирования экспрессии ядерного генома. Эти взаимодействия детерминируют широкий спектр различных фенотипических эффектов на протяжении всего продукционного процесса (особенности роста и развития, устойчивость и др.), что обеспечивает более высокий уровень адаптации растений к стрессовым факторам среды.